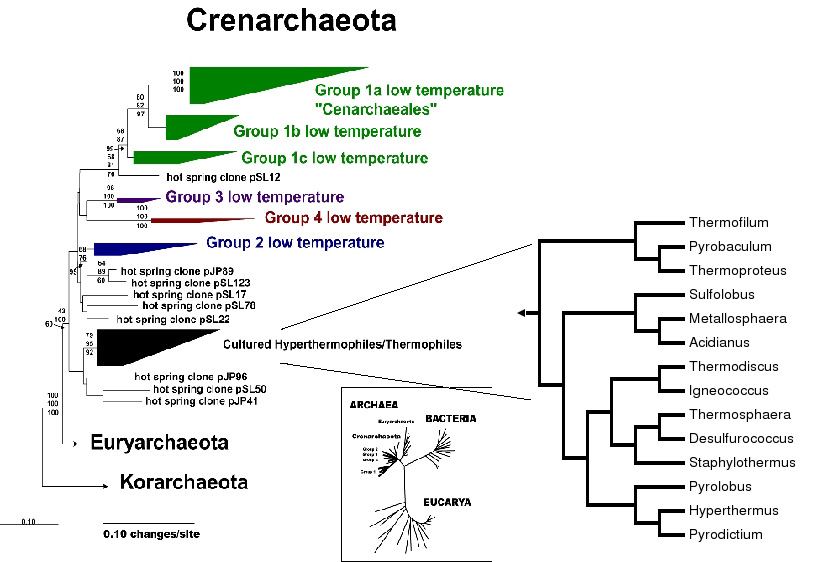

Possible metabolism of low temp crens: Groups 2, 3, and 4 problably are involved in sulfur chemistry. Group 1, which is the most abundant and wide spread is likely aerobic and likely constitute a significant component, about 30 percent, of the microbes in the deep sea.
Cultured Crens that use sulfur:
H2 and sulfur in anoxic media producing sulfide were used to enrich novel extremely thermophilic archaea were reported by Fischer et al., 1983; Stetter, 1982; 1983a; 1983b; Zillig et al., 1981; 1982; 1983, and the number of novel isolates increased steadily in subsequent years (for overview see e.g., Stetter et al., 1990; 1996). Several isolates seemed to reduce sulfur in a by-reaction or as mere electron sink to facilitate fermentation (Zillig et al., 1982; for more recent overview see Schönheit and Schäfer, 1995; Hedderich et al., 1999). Nevertheless, evidence for sulfur respiration as a mode of energy metabolism in archaea was clearly provided in cultures of Thermoproteus and Pyrodictium species that grew with H2 as the only electron donor in the absence of organic compounds (Fischer et al., 1983; Stetter et al., 1983b). Further, newly isolated archaea that definitely grow by sulfur respiration, viz on H2 and sulfur, were Stygioglobus azoricus (Segerer et al., 1991), Pyrobaculum islandicum (Huber et al., 1987) and Stetteria hydrogenophila (Jochimsen et al., 1997).
A unique versatility in sulfur metabolism was found in new lithoautotrophic thermophilic isolates, Acidianus infernus (Segerer et al., 1985; Segerer et al., 1986) and Desulfurolobus (originally Sulfolobus) ambivalens (Zillig et al., 1985; Zillig et al., 1986) that grew aerobically by sulfur oxidation as well as anaerobically by sulfur reduction with H2.
In carbon assimilation during sulfur reduction with H2, the reductive citric acid cycle and more recently the hydroxypropionate pathway were shown to be operative in Thermoproteus neutrophilus (Schäfer et al., 1986) and Acidianus (Menendez et al., 1999), respectively. In the course of investigations into the sugar metabolism in several hyperthermophiles (for overview see Selig et al., 1997), pathways also were investigated in the sulfur-respiring, facultatively organotrophic Thermoproteus tenax (Siebers and Hensel, 1993). Evidence was provided for a non-phosphorylated Entner-Doudoroff pathway and a modified Embden-Meyerhof-pathway. Furthermore, complete oxidation of organic substrates via the citric acid cycle was demonstrated in the facultatively organotrophic sulfur-respiring species Thermoproteus tenax and Pyrobaculum islandicum (Selig and Schönheit, 1994). So far, these are the only extremely thermophilic sulfur-reducing microorganisms shown to couple sulfur reduction to complete mineralization of organic compounds, analogous to Desulfuromonas and Desulfurella (see above). The electron transport during sulfur reduction with H2 was studied in Pyrodictium brockii (Phil et al., 1992; Maier, 1996) and Pyrodictium abyssii (Dirmeier et al., 1998); these species employ different transport chains.